
科学网—清华大学方晓峰团队发现细胞水势感受器
已有 10718 次阅读
2026-5-30 04:08
| 个人分类: 科研思路分享 | 系统分类: 科研笔记
清华大学方晓峰团队发现细胞水势感受器
水是生命的最基本要素。对于植物而言,种子干燥,干旱、高盐和高温等逆境胁迫,最终都会归结为同一个核心问题 — 细胞内「可利用水」变少。长期以来,人们已经认识到细胞能够感知渗透压变化,并通过一系列信号级联通路启动应答。然而,一个更基础的问题始终缺乏答案:细胞究竟能否直接感知「水本身」的变化?
2026 年 5 月 28 日,清华大学方晓峰团队在 Nature 杂志在线发表了题为 Cellular water potential sensing via biomolecular condensation 的研究论文,首次揭示植物细胞能够通过一种「水势依赖的生物大分子相分离」机制,直接感知细胞水状态变化,并进一步重塑 RNA 转运与翻译过程,从而促进植物适应逆境环境。
Nature杂志同期以 Research Briefing 的形式对该工作进行了题为「Troubled waters: a protein senses when cells are running dry」的报道。
基于生物分子凝聚的细胞水势感知
水分子作为生物大分子的溶剂,对细胞至关重要。在缺水条件下,细胞水势会降低,然而细胞如何感知水势变化仍不清楚。本研究鉴定出一种含无菌 α结构域(SAM)的蛋白SAM8,该蛋白在体内和体外均会发生水势依赖性凝聚,对高渗胁迫耐受与种子萌发至关重要。我们利用生物物理技术、体外重组与生物成像证明:正常水分条件下SAM8高度水合,阻止其发生宏观凝聚;带负电荷的区域通过构建电场与微极性环境决定SAM8的水合状态。缺水条件会削弱这种水合作用,进而通过重编程氢键、静电相互作用与疏水相互作用激活SAM8凝聚。此外,我们证实SAM8凝聚体可选择性截留RNA输出因子,导致高渗胁迫下mRNA滞留于细胞核并引发翻译重编程。本研究揭示了植物细胞直接感知并响应水分状态的机制,为阐明植物如何适应缺水环境提供了新见解。
---
正文
水分子是生命的核心,为细胞内生物大分子维持功能结构与活性提供溶剂环境。细胞内的水分子与大分子结合形成水合层,或在本体中自由扩散,这两部分水分别被称为界面水与自由水。水势可理解为自由水的可利用度,决定植物从土壤中吸水以及体内水分运输。细胞水势对环境波动十分敏感,尤其是干旱、高盐与温度胁迫。活细胞内的生物大分子至少被一层分子厚度的水合水包裹。细胞水势降低会导致生物大分子水合度下降,破坏膜完整性、扰乱蛋白质三维结构、削弱酶活性等。因此,细胞水势的感知与响应对植物发育及适应各类环境胁迫至关重要,但其背后机制仍不明确。
SAM8发生水分依赖性凝聚
生物分子凝聚正成为感知与响应环境胁迫的关键机制。高渗胁迫常引发细胞体积收缩,导致膜张力改变、水势降低,进而使生物大分子脱水、分子拥挤度与离子浓度上升。已有研究报道,植物中细胞核与细胞质蛋白可通过分子拥挤依赖性凝聚感知高渗胁迫。我们推测,生物分子凝聚可作为感知细胞水势变化的方式,因为水分可调控疏水相互作用、静电相互作用或阳离子 –π相互作用的强度,而这些均是凝聚的驱动力。重水(D₂O)形成的氢键比普通水(H₂O)更强,会增强水–水相互作用并减弱水–蛋白相互作用。因此,我们使用重水处理,在不改变细胞体积的前提下减少与蛋白结合的水分。
图 1
a, Illustration of the impact of osmotic stress and D2O on plant cells. b, Representative confocal images of tobacco epidermal cells (top) or Schizosaccharomyces pombe cells (bottom) expressing SAM8-GFP. The cells were treated for 10 min with an isosmotic buffer prepared with H2O or 90% D2O. The white dotted circles indicate the nuclei of yeast cells. Scale bar, 5 μm. c, Quantification of the percentage of cells containing SAM8 condensates as indicated. Error bars indicate mean ± s.d. (n = 10 independent experiments). d, Representative confocal images of Arabidopsis thaliana root tip cells that were treated with an isosmotic buffer prepared with H2O or 90% D2O. Scale bar, 5 μm. e,f, Representative confocal images of Arabidopsis root tip cells expressing SAM8-mVenus. The cells were treated with 0.3 M mannitol or 0.5 M EG for 10 min. Scale bar, 5 µm. g, Illustration of the dehydration and hydration process during seed development and germination. h, Representative confocal images of embryos dissected from the different stages of seeds expressing SAM8. DAP, days after pollination. Scale bar, 5 μm. i, Representative confocal images of embryos dissected from the dry and imbibed seeds expressing SAM8. Scale bar, 10 μm (white); 2 μm (yellow). Representative images of n independent experiments (n = 3 (d–f, h and i)). Asterisks indicate significant differences (two-tailed t-test).
为阐明植物细胞如何直接感知水势,我们筛选具有相分离潜能的蛋白,在细胞中表达后用重水配制的等渗缓冲液处理,寻找重水依赖性凝聚蛋白。通过该筛选,我们在烟草细胞中发现数个可响应重水处理形成凝聚体的蛋白。其中, At5g23680编码一种含SAM结构域的蛋白,命名为SAM8,重水处理可诱导其在烟草与酵母细胞中形成细胞核凝聚体。我们随后在sam8-1敲除植株中构建SAM8互补株系(SAM8pro::SAM8-mVenus)。在根尖细胞中,SAM8-mVenus弥散分布于核质;将植株转入含40%以上重水的等渗缓冲液处理5分钟,SAM8即形成细胞核凝聚体。免疫染色结果也证实了重水诱导的SAM8凝聚。作为对照,两种已知的相分离蛋白SEUSS与DCP5在重水处理下均不形成凝聚体,上述结果表明SAM8对重水的响应真实且特异。
我们进一步验证 SAM8凝聚是否响应高渗胁迫诱导的水势变化。高渗胁迫处理后,SAM8在烟草表皮细胞、酵母细胞与拟南芥根尖细胞中均快速形成凝聚体。SAM8发生凝聚的水势变化阈值约为0.3–0.35 MPa。该凝聚体具有高度动态性,胁迫解除后20分钟内即可解聚,光漂白后荧光也能快速恢复。乙二醇(EG)不会引发分子拥挤但可降低细胞水势,同样能触发SAM8凝聚,说明SAM8凝聚不依赖分子拥挤。渗透信号早期信号——脱落酸(ABA)与活性氧(H₂O₂)处理均无法诱导SAM8凝聚。此外,LaCl₃化学抑制钙内流,或Raf样激酶基因敲除(ok130-null突变体),对SAM8凝聚几乎无影响。这些结果表明,SAM8凝聚不依赖已知的渗透信号通路。
种子成熟与萌发过程中细胞水势发生显著变化。 SAM8在种子与根组织中高表达,促使我们探究其在种子发育与萌发中的凝聚行为。种子发育早期胚胎水分充足时,SAM8在细胞核内弥散分布;伴随发育后期脱水与种子干燥,SAM8形成凝聚体。种子在水中吸胀后,SAM8凝聚体在10–20分钟内快速解聚,而在NaCl溶液中吸胀则不解聚;将水中吸胀的种子转入NaCl溶液,SAM8凝聚体重新出现,说明SAM8凝聚可动态响应种子内的水势变化。
温度也可调控细胞水势。已有研究表明,温度升高与降低分别拮抗与协同高渗处理对细胞水势的影响。我们发现, 35℃下的高渗处理诱导的SAM8凝聚体少于25℃,而15℃则显著增强酵母细胞中SAM8的凝聚。统计分析显示,外界渗透强度与温度对SAM8凝聚存在显著交互作用,说明温度与渗透胁迫共同调控SAM8凝聚。同样,在拟南芥根中,25℃下高渗处理5分钟即可触发SAM8凝聚,而37℃下相同渗透强度处理20分钟仍无法诱导凝聚。
综上,这些数据证明 SAM8在细胞内发生水势依赖性凝聚。
SAM8凝聚的驱动力
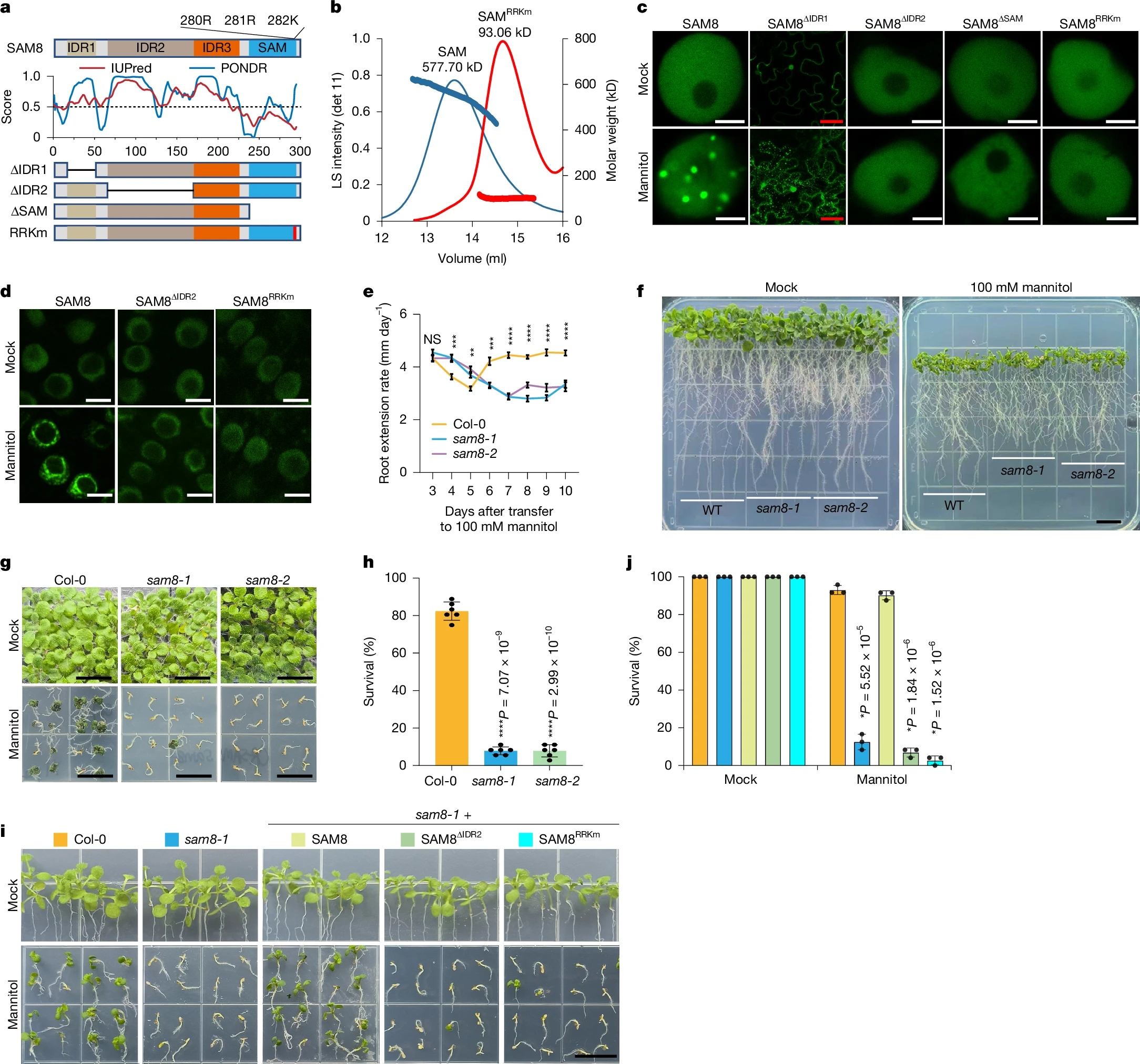
SAM8的C端含有一个SAM结构域,其余区域大多为非折叠状态,被预测为内在无序区(IDR)。SAM结构域是经典的寡聚化结构域,可发生头尾组装,已有报道其介导相分离。基于序列的预测显示,SAM结构域末端螺旋表面的三个带正电氨基酸(280R、281R、282K)对SAM8寡聚化至关重要。体外纯化的SAM结构域主要以寡聚体形式存在,而将这三个RRK残基突变为带负电的谷氨酸残基(SAMRRKm)后,SAM转变为单体。为探究各区域对SAM8凝聚的贡献,我们构建截短或突变变体并在烟草细胞中检测其凝聚行为。缺失IDR1会使SAM8部分分布于细胞质,但仍保留渗透依赖性凝聚;缺失IDR2则完全丧失凝聚能力。删除SAM结构域(SAM8∆SAM)或突变SAM结构域(SAM8RRKm)的SAM8均无法形成凝聚体。与此一致,共表达SAM8∆IDR2与SAM8RRKm可重建渗透依赖性凝聚。酵母细胞实验进一步证实IDR2与SAM结构域对SAM8凝聚的必要性。在拟南芥稳定转基因植株中,SAM8∆IDR2与SAM8RRKm在甘露醇或重水处理下均无法形成凝聚体。这些结果表明,IDR介导的多价相互作用与SAM介导的寡聚化均为SAM8凝聚所必需。
图2
a, Top and bottom, domain structures of SAM8 and its variants. IDR, intrinsically disordered region. Middle, IDR prediction by the indicated algorithms. b, Size-exclusion chromatography coupled with multi-angle laser light scattering assay showing the size of the SAM domain and SAMRRKm. Both proteins were fused with an MBP tag to prevent misfolding. Left y-axis, the light scattering intensity from the 90° detector (detector 11). Right y-axis, molecular weight. c, Representative confocal images of tobacco epidermal cells expressing SAM8 and its variants. The cells were treated with or without 0.3 M mannitol. Scale bars, 5 µm (white); 50 µm (red). d, Representative confocal images of Arabidopsis root tip cells expressing SAM8 and variants treated with or without 0.3 M mannitol for 10 min. Scale bar, 5 µm. e, Quantification of the root extension rate of Arabidopsis seedlings that were transferred to medium supplemented with 100 mM mannitol. Error bars indicate mean ± s.e. (n = 30). Statistical significance details are provided as Source Data. f, Representative images of Arabidopsis seedlings that grew for 10 days after transferring to medium supplemented with or without 100 mM mannitol. Scale bar, 1 cm. g,i, Representative images of 12-day-old seedlings of the indicated genotypes grown on medium containing 300 mM mannitol. Scale bar, 1 cm. h,j, Quantification of the survival percentage shown in g and i, respectively. Error bars indicate mean ± s.d. (n = 6 replicates with each replicate containing 36 plants for h; n = 3 with each replicate containing approximately 24 plants for j). For e,h and j, asterisks indicate significant differences (two-tailed t-test). NS, not significant. For c and d, n = 3 independent experiments.
SAM8凝聚具有生理功能
我们随后评估 SAM8凝聚的功能意义。获得两个突变等位基因:sam8-1为T-DNA插入突变体,SAM8表达完全缺失;sam8-2为CRISPR突变体,1 bp插入导致提前终止密码子。正常生长条件下,sam8突变体与野生型Col-0表型无差异。植株暴露于高渗胁迫时,Col-0的根尖分生组织生长与根伸长速率在最初几天受抑,随后部分恢复;而sam8突变体的这种抑制与恢复能力显著受损。据此推测,渗透胁迫下生长抑制缺陷会增加胁迫敏感性并可能引发细胞死亡。SYTOX Green染色显示,sam8突变体的细胞死亡率显著高于Col-0,高渗胁迫下sam8幼苗的存活率显著降低。野生型SAM8可完全回补sam8-1突变体的高渗敏感表型,而凝聚缺陷变体则无法回补。这些数据证明,SAM8凝聚对植物渗透胁迫适应与耐受至关重要。
自然界中,高渗胁迫常与环境温度升高或热胁迫同时发生。 SAM8凝聚同时依赖渗透势与温度,促使我们探究其是否介导二者的协同效应。结果显示,26℃下的高渗处理相比22℃,显著提高野生型植株的死亡率。sam8-1突变体在两种温度下的渗透处理中根生长均极弱。与此一致,在两种温度的高渗培养基上萌发并生长时,大部分Col-0幼苗在22℃存活,而26℃下约35%死亡;sam8突变体在22℃与26℃下无差异,SAM8互补植株则与Col-0表型一致。相反,SAM8过表达提升凝聚倾向,增强植株在22℃与26℃下的存活率。因此,SAM8是通过水势依赖性凝聚整合渗透胁迫与温度信号的胞内模块之一。
鉴于 SAM8凝聚也响应种子中的水势变化,我们探究其在种子萌发中的功能。结果显示,sam8-1与sam8-2的萌发速率均晚于Col-0,该缺陷可被野生型SAM8完全回补,而凝聚缺陷变体则无法回补。综上,这些数据证明SAM8的水势依赖性凝聚具有生理功能。
SAM8在体外响应水势变化
为阐明水势依赖性凝聚的分子机制,我们纯化 SAM8蛋白并检测其体外相行为。SAM8-GFP蛋白均匀弥散,加入PEG8000后形成球形液滴;而相分离实验中常用的分子拥挤剂葡聚糖(dextran)或聚蔗糖(ficoll)则无法触发SAM8相分离。我们使用已报道的分子拥挤传感器SEU的IDR1验证该结果:4% PEG与7.5%葡聚糖均可诱导IDR1SEU相分离,但只有4% PEG可诱导SAM8相分离。
图 3:SAM8感知水势的分子基础
本图替代文本可能由 AI生成。
a,在所示拥挤剂存在下,对2.5 μM SAM8进行体外相分离实验。
b,蒸汽压渗透法测量渗透势(n=5次独立实验)。
c,水合半径(Rh)与回转半径(Rg)示意图。
d,SAM8的Rg与Rh测定(DLS n=6;SLS n=3)。
e,DLS测定所示溶液的水合半径。
f,上:IDR3或IDR3mQ的氨基酸序列;下:CIDER预测的SAM8各残基净电荷。
g,SAM8IDR3mQ的Rh与Rg测定(DLS n=30;SLS n=3)。
h,2.5 μM SAM8与SAM8IDR3mQ的体外相分离实验。
i,FLIM-FRET实验中供体与受体示意图。
j、k,PEG 8000(j)或葡聚糖(k)存在下供体的荧光寿命(n=20个事件)。
l,所示蛋白的GFP供体荧光寿命(n=30个事件)。
m,DI-4-ANEPPS染料存在下5 μM SAM8与SAM8IDR3mQ的共聚焦成像。
n,m中535–545 nm与610–640 nm发射光的比值(n=98个凝聚体)。
o,双SBD标记的SAM8与SAM8IDR3m凝聚体的FLIM成像。
p,o中双SBD荧光寿命定量(n=9个独立样本)。
*表示差异显著(双侧t检验)。NS:无显著差异。
q,在sam8突变体背景中表达的SAM8与SAM8IDR3mQ的成像。
误差线表示均值 ±标准差。 *p<0.05 ,双侧 t检验。NS:无显著差异。
比例尺: 10 μm(a、m、o);5 μm(h、q)。
a、e、h、m、o、q:n=3次独立实验。
尽管 PEG与葡聚糖诱导分子拥挤的能力相近,但PEG能够结合远更多的水分子,从而显著降低水势。我们通过蒸汽压渗透法证实了这一点(图3b)。SAM8对PEG而非葡聚糖敏感,说明SAM8在体外的相分离依赖水势。与此一致,SAM8-GFP在重水配制的缓冲液中形成凝聚体,而在普通水中则不会(扩展数据图5d);能够强力剥离生物分子表面水合水的小分子三氟乙醇(TFE),也可诱导SAM8凝聚(扩展数据图5e)。
GFP标签不影响SAM8凝聚,因为不带标签的SAM8在加入重水或PEG后同样形成凝聚体(扩展数据图5d、f)。作为对照,SEUSS-IDR1、DCP5-ICS、FREE1的凝聚不受重水影响(扩展数据图5d)。与体内观察一致,SAM8在体外发生相分离的能力主要依赖IDR2与SAM结构域(扩展数据图5g、h)。这些数据表明,SAM8的凝聚本质上对水势下降敏感。
SAM8具有厚水合层
为理解水势响应的分子基础,我们研究了 SAM8的水合状态。我们利用动态光散射与多角度光散射分别测定SAM8的水合半径(Rh)与实际尺寸(回转半径,Rg)(图3c)。我们发现SAM8的Rh显著大于Rg(图3d),说明其处于高度水合与舒展构象。作为对照,同样含有聚合结构域的SOSEKI1与SAM7的Rh和Rg无明显差异(扩展数据图5i)。与PEG争夺水合水的能力一致,加入PEG后SAM8的Rh下降(图3e)。高温下PEG对水势的影响减弱(扩展数据图5j、k)。我们发现,相同PEG条件下,37℃时SAM8液滴的尺寸显著小于25℃(扩展数据图5l、m)。相比之下,IDR1SEU的液滴在两种温度下无明显变化(扩展数据图5l、m)。综上,这些结果证明SAM8具有厚水合层,水势降低可促使其在体外发生相分离。
水势感知的分子基础
我们接下来探究 SAM8水合的机制。净电荷分析显示,SAM8的IDR3区域高度带负电(图3f,扩展数据图6a)。基于AMOEBA力场的能量计算显示,天冬氨酸Asp与谷氨酸Glu具有极低的固有水合自由能。因此我们提出:IDR3中的负电荷是SAM8水合的主要决定因素。为验证这一点,我们构建了SAM8突变体(SAM8IDR3mQ),将IDR3中的Asp、Glu分别突变为Asn、Gln(图3f)。与野生型SAM8不同,SAM8IDR3mQ的Rh与Rg无差异(图3g),且无需PEG即可形成凝聚体(图3h)。
为区分静电排斥与水合作用对 SAM8凝聚的贡献,我们将IDR3中10个核心Asp/Glu突变为丝氨酸(SAM8IDR3mS;扩展数据图6b、c),以保留水合作用但消除静电排斥——因为丝氨酸具有极性,可通过偶极-偶极作用吸引水分子。我们还构建了SAM8IDR3mK与SAM8IDR3mR(扩展数据图6b、c),以保留静电排斥但削弱水合作用——因为Lys、Arg的水合能力弱于Asp、Glu。生物物理分析证实,SAM8IDR3mS的Rh与Rg仍存在显著差异,而SAM8IDR3mK与SAM8IDR3mR的差异很小或无差异(扩展数据图6d)。与此一致,SAM8IDR3mS仍以PEG依赖的方式形成凝聚体,而SAM8IDR3mK与SAM8IDR3mR无需PEG即可组成型凝聚(扩展数据图6e)。这些结果表明:带负电的IDR3主要通过强水合作用控制SAM8凝聚。
随后我们进行 FLIM-FRET实验,检测水势变化下SAM8的分子间相互作用(图3i)。使用SAM8∆SAM以避免SAM寡聚化对FRET信号的主导影响。结果显示,随PEG浓度升高,SAM8分子间相互作用逐渐增强(图3j);NaCl(扩展数据图6f、g)、甘露醇(扩展数据图6h、i)也有类似效果,但葡聚糖无此作用(图3k)。值得注意的是,SAM8在体外发生相互作用的水势阈值(图3b)与体内凝聚阈值(扩展数据图1i、j)相似,说明SAM8可感知生理相关的水势阈值。破坏IDR3的负电荷显著增强分子间相互作用(图3l)。综上,这些数据证明:水势降低直接促进SAM8自互作并触发凝聚。
鉴于负电氨基酸具有强电荷 -偶极作用与氢键能力,我们接下来检验负电荷是否在SAM8周围形成电场,从而极化并束缚水分子(图3m、n)。为此我们检测SAM8凝聚体的微环境。Di-4-ANEPPS可用于测量局部电场。我们发现,SAM8凝聚体的ANEPPS信号比值显著高于SAM8IDR3mQ凝聚体(图3m、n)及其他突变体(扩展数据图6j、k),说明SAM8凝聚体内电场更强。SBD类荧光团可用于测量凝聚体微极性:极性越高,荧光寿命越短。SAM8凝聚体的荧光寿命短于SAM8IDR3mQ凝聚体(图3o、p)。这些结果共同表明:IDR3中的负电荷赋予SAM8强水合,在水分充足时阻止其凝聚。
SAM8凝聚的保守性分析
我们进行进化分析,在所有陆生植物中均鉴定到 SAM8同源蛋白(扩展数据图7a)。这些同源蛋白均含有高度保守的C端SAM结构域,旁侧为保守的负电区域(扩展数据图7a)。值得注意的是,不同SAM8同源物的负电程度不同,我们推测这与其凝聚所需的水势阈值相关。我们发现,耐旱作物亚麻荠(Camelina sativa,Cs)的SAM8与拟南芥SAM8高度同源,但其IDR3的负电氨基酸序列更长(扩展数据图7b)。体外实验显示,CsSAM8需要略高浓度的PEG才能发生相分离(扩展数据图7c)。
在酵母细胞中,随山梨醇浓度升高, AtSAM8与CsSAM8均形成更多凝聚体,支持SAM8响应水势的保守性。但相同处理下,CsSAM8形成的凝聚体更少(扩展数据图7d),说明CsSAM8的水势响应阈值更低。随后我们在sam8-1突变体中回补表达CsSAM8。与AtSAM8相比,相同甘露醇或重水处理下,CsSAM8形成的凝聚体更少(扩展数据图7e)。根伸长实验显示,相同渗透胁迫下,CsSAM8回补植株受抑制程度更轻(扩展数据图7f–i)。与此一致,SAM8IDR3mQ在正常条件下即可在根尖细胞核组成型形成凝聚体(图3q),且SAM8IDR3mQ回补植株生长发育迟缓(扩展数据图7j)。综上,这些结果表明:SAM8感知水势的能力在进化上保守,对平衡植物生长与渗透胁迫耐受至关重要。
图 4:SAM8凝聚体选择性招募RNA输出因子
a,RNA输出关键步骤示意图。
b,烟草表皮细胞中共表达SAM8与ALY的Co-IP实验。
c,烟草细胞核中ALY-GFP与SAM8-Cerulean凝聚体共定位。比例尺:5 μm。
d,体外相分离显示ALY1与SAM8液滴共定位。比例尺:5 μm。
e,2.5 μM ALY1-BFP与UAP56B-mCherry在递增浓度SAM8-GFP下的体外相分离。比例尺:5 μm。
f,e中液滴内部平均荧光强度。误差线:均值±标准差,n=6。 *p<0.05 ,双侧 t检验。
g,烟草细胞共表达显示SAM8凝聚体招募ALY1与EIF4A3,但排斥UAP56B。比例尺:5 μm。
b–d、g:n=3次独立实验。
细胞核 mRNA输出是保证基因正常表达的关键RNA监控步骤。近期人类mRNA输出复合物结构解析显示,ALYREF(ALY)通过外显子连接复合物(EJC)识别mRNA核糖核蛋白复合物(mRNP),并通过UAP56连接转录-输出复合物(TREX),形成可输出的TREX-mRNP复合物(图4a)。为阐明SAM8凝聚的分子功能,我们在高渗胁迫下通过免疫沉淀寻找SAM8互作蛋白。GO分析显示mRNA监控与RNA剪接显著富集(扩展数据图8b)。值得注意的是,互作组中鉴定到多个RNA输出因子,包括ALY家族全部4个成员与EJC亚基EIF4A3(扩展数据图8a,附表2)。SAM8与ALY、EIF4A3的相互作用通过Co-IP(图4b)、BiFC(扩展数据图8c)与FLIM-FRET(扩展数据图8d)验证。
在烟草表皮细胞中, ALY与EIF4A3在核仁与核质中弥散分布,与拟南芥中的定位一致(图4c)。高渗处理后,ALY与EIF4A3高度富集于SAM8凝聚体(图4c),说明SAM8凝聚体可分配ALY与EIF4A3。类似地,ALY与EIF4A3在体外也被SAM8液滴招募(图4d)。
ALY家族蛋白具有保守的中央RRM结构域,可非特异性结合单链RNA(扩展数据图8e)。我们发现ALY1可与35 nt PolyA RNA共同形成液滴。将ALY/RNA与SAM8共孵育时,ALY/RNA液滴在SAM8液滴内部形成亚相(扩展数据图8f),类似核仁蛋白形成的多相共存结构。ALY-RNA亚相的形成不受添加顺序与孵育时间影响(扩展数据图8g、h),可能源于SAM8凝聚体更高的微极性,或SAM8、ALY、RNA之间相互作用强度差异。这些结果进一步支持SAM8凝聚体分配ALY/RNA复合物。
拟南芥 ALY可直接结合UAP56。与此一致,UAP56在体外可被招募进ALY1液滴(图4e)。但UAP56未出现在SAM8免疫沉淀中(附表1),也不进入SAM8凝聚体(扩展数据图8i)。向ALY1/UAP56中加入递增浓度SAM8后,只有ALY1被分配进入SAM8液滴,UAP56B则不能(图4e、f)。烟草细胞共表达证实,SAM8凝聚体招募ALY1与EIF4A3,但排斥UAP56(图4g)。这些发现表明:SAM8凝聚体可通过选择性分配破坏ALY与UAP56的结合。
图 5:SAM8介导渗透胁迫下mRNA核滞留与翻译重编程
a,拟南芥根尖oligo(dT)探针RNA FISH。比例尺:5 μm。
b,a中mRNA核质比定量。误差线:均值±标准差。 *p<0.05 ,单因素方差分析。 NS:无显著差异。
c,300 mM甘露醇上生长12天的幼苗表型。比例尺:1 cm。
d,c中存活率统计。误差线:均值±标准差。
e、f,甘露醇处理下Col-0(e)与sam8-1(f)的翻译效率(TE)变化。
g,Col-0与sam8-1中TE变化基因的重叠。
h,甘露醇处理下sam8-1 vs Col-0的TE上调/下调基因前20个GO条目。
i,多聚核糖体分析验证候选基因TE。误差线:均值±标准差,n=3。
j,SAM8工作模型。水分充足时SAM8高度水合,不凝聚;缺水时水合减弱,SAM8凝聚;SAM8凝聚体选择性招募/排斥RNA输出因子,阻断mRNA输出,引发翻译重编程。
j图由BioRender绘制。
SAM8凝聚体选择性招募RNA输出因子,促使我们提出:SAM8凝聚可抑制mRNA运输。为验证这一点,我们用oligo(dT)探针进行FISH检测mRNA定位。正常条件下,野生型Col-0的mRNA在细胞核与细胞质均匀分布(图5a、b);渗透处理后,mRNA核质比显著升高(图5a、b)。相比之下,sam8-1突变体在正常与胁迫下核质比无明显变化(图5a、b)。这些发现表明:渗透胁迫以SAM8凝聚依赖的方式引发mRNA核滞留。
为检验 mRNA核滞留对渗透适应的重要性,我们过表达ALY与EIF4A3以增强mRNA输出。结果显示,这导致植株对渗透胁迫更敏感(图5c、d,扩展数据图9a–c)。这些数据表明:SAM8凝聚依赖的RNA输出调控促进渗透耐受。
为检验这种滞留如何调控翻译,我们对 Col-0、sam8-1、sam8-2在正常与高渗条件下同时进行mRNA-seq与Ribo-seq,计算全局翻译效率(TE)。质控见扩展数据图10a、b。结果显示,高渗在Col-0中引发显著的TE变化(图5e)。GO分析显示,TE上调基因富集于胁迫响应,TE下调基因富集于生长发育(扩展数据图10c),说明植物通过翻译重编程平衡生长与防御。相比之下,sam8突变体中高渗诱导的TE变化小得多(图5f,扩展数据图10d),与野生型重叠不足1%(图5g),支持SAM8对渗透胁迫响应的翻译重编程至关重要。
比较 sam8-1与野生型的翻译效率,在对照与胁迫下均出现显著变化(扩展数据图10e、f)。GO分析显示这些基因主要富集于发育与胁迫相关过程(图5h,扩展数据图10g),说明SAM8参与平衡生长与胁迫防御。多聚核糖体分析验证候选基因翻译(扩展数据图10h)。与Ribo-seq一致,渗透胁迫下Col-0中胁迫关键因子ABF4、DREB2A的翻译上调,而sam8-1中不上调(图5i);根分生组织关键调控因子LHW、RGI4的翻译在Col-0中显著下调,但在sam8-1中基本不变(图5i)。作为对照,UBC10的翻译效率不受影响(扩展数据图10i)。
综上,这些结果表明:渗透胁迫下 SAM8凝聚通过调控mRNA输出,促进胁迫适应型翻译并抑制生长相关翻译。
---
讨论 Discussion
本研究结果共同证实, SAM8 是一个细胞水势直接感受器。SAM8 具有由 SAM 结构域介导的寡聚化作用,以及由内在无序区(IDR)介导的弱多价相互作用,使其易于发生凝聚。然而,SAM8 内部的一段高负电区域能够形成电场,从而维持蛋白表面的厚水合层。
因此,在细胞水分充足的条件下, SAM8 的高度水合状态会阻止其发生凝聚;而当植物遭遇干旱、高盐等水分缺失环境时,细胞水势下降,SAM8 水合作用减弱,进而触发 SAM8 发生相分离形成凝聚体(图 5j)。SAM8 凝聚体能够选择性招募与排斥 RNA 输出因子,导致 mRNA 核滞留,并最终引发翻译重编程(图 5j)。
尽管生物分子凝聚的形成通常受水 —蛋白相互作用影响,但蛋白—蛋白相互作用同样是构成生物分子凝聚热力学驱动力的重要部分。SAM8 的独特性在于:它同时具备由 SAM 结构域介导的强蛋白—蛋白相互作用,以及由负电区域介导的强蛋白—水相互作用。我们认为,蛋白—蛋白相互作用与蛋白—水相互作用的组合与平衡,是实现水势感知的关键。许多其他聚合结构域与疏水区域也参与生物分子凝聚,本发现为新型水势感受器的发现与人工设计提供了新思路。
面对多种胁迫时,真核细胞会激活整合应激反应( ISR),其核心是减弱全局蛋白合成,同时促进特定 mRNA 的选择性翻译。已有研究表明,高渗胁迫会抑制植物中大量 mRNA 的翻译。RNA 输出是保证转录与翻译连续性的关键环节。本研究结果表明,SAM8 凝聚通过控制 mRNA 输出实现翻译重编程,代表了植物在高渗胁迫下调整翻译的重要策略之一。
已有研究报道,在酿酒酵母中, RNA 输出可作为热胁迫下的监控机制:高温促使 RNA 输出因子 Mex67 从常规 mRNA 上解离,阻止普通 mRNA 输出;而热激 mRNA 则可快速输出并优先翻译,以提升细胞在极端条件下的存活率。众所周知,胁迫抗性与生长之间的平衡受到精细调控,SAM8 凝聚可能正是维持这种平衡的关键机制之一。我们未来的研究方向是:检验植物中是否存在某些胁迫相关 mRNA 能够逃逸 SAM8 介导的核滞留,并被选择性输出到细胞质中。
---
方法 Methods
植物材料、培养条件与胁迫处理
本研究中所有拟南芥突变体与转基因植株均为 Columbia(Col-0)背景。T-DNA 插入突变体 sam8-1(SALK_065676)购自 AraShare。sam8-2 是 CRISPR 编辑突变体,在起始密码子下游 39 bp 处发生 1 bp 插入,导致移码突变与提前终止,其编码蛋白序列为 MAELQLVEGHQINRRFYPAGDNKLNRSTGNIRRSRSFSRIETIEKT*。ok130 敲除突变体由南方科技大学王鹏程教授惠赠。
种子用 2.5%(v/v)次氯酸钠与 70%(v/v)乙醇表面消毒,4℃黑暗春化 3 天,点播于 1/2 MS 固体培养基(含 1% 蔗糖、0.8% 琼脂)。培养条件为长日照:16 小时光照(22℃)/8 小时黑暗(18℃)。
萌发动力学实验
所有基因型种子同期采收、相同条件栽培。 37℃均匀干燥 2 周以保证一致的后熟过程。随后点播于同一批培养基,长日照培养(22℃光照/18℃黑暗),不春化。以种皮破裂为萌发标准计算萌发率。每个基因型设 3 次生物学重复,每重复约 300 粒种子。
直接渗透胁迫处理
消毒后的种子点播于含 300 mM 甘露醇的 1/2 MS 培养基,分别在 22℃ 或 26℃ 培养,12 天后记录表型。存活率判定:胁迫解除后幼苗能否恢复生长。具体为胁迫处理后转回正常 1/2 MS 培养基,7 天后统计:保持绿色并长出新组织(真叶)为存活;变白/变褐无法恢复为死亡。
移栽实验
Col-0 与 sam8-1 种子在正常 1/2 MS 培养基、22℃ 萌发 3 天,移栽至含 750 mM 甘露醇的培养基,22℃ 或 26℃ 垂直培养 5 天,处理结束后移栽回正常培养基,22℃ 培养 10 天后记录表型。
根伸长实验
5 天龄幼苗移栽至含甘露醇的培养基,垂直培养 10 天。分生区长度测量:移栽 2 天后使用 Nikon AXR 共聚焦显微镜 100× 油镜采集微分干涉图像,从静止中心(QC)量至第一个伸长细胞。根伸长速率:每日测量根长,以 mm/天计算。
质粒构建
构建 pSAM8::SAM8-mVenus:从野生型基因组扩增 2.2 kb 启动子与 1 kb 3′UTR,连接到 pCAMBIA1300-mVenus 载体,得到 pSAM8::mVenus-UTR;再将 SAM8 编码序列插入启动子与 mVenus 之间。
SAM8 突变体与截短体:通过定点突变或结构域缺失 PCR 构建 SAM8ΔIDR1、SAM8ΔIDR2、SAM8IDR3mQ、SAM8IDR3mS、SAM8IDR3mK、SAM8IDR3mR、SAM8ΔSAM、SAM8RRKm,测序验证后插入上述载体。
构建 sam8-2:设计两个 sgRNA 插入 pAtU6-26-M 载体,Cas9 表达框亚克隆至 pCambia1300-UBQ:Cas9-P2A-GFP 载体。所有载体通过农杆菌 GV3101 转化,花侵染法转化 sam8-1,潮霉素筛选。纯合株系用于实验;sam8-2 使用 Cas9-free 株系。
烟草瞬时表达载体: SAM8 及其突变体、ALY1/2/3/4、eIF4A3、UAP56B、NUL1 扩增后插入 pCAMBIA1300-35S 系列荧光标签载体。
酵母异源表达载体: SAM8 编码序列插入 pDUAL-Pnmt1-yeGFP。
体外蛋白表达载体: SAM8 及其突变体、ALY1–4、eIF4A3、UAP56B、SAM7、SOSEKI1 插入 pET11 系列带 His/GFP/mCherry/BFP 标签载体。部分载体 N 端融合 MBP 可溶性标签与 TEV 酶切位点。极性检测载体:MBP-SAM8 / MBP-SAM8IDR3mQ 插入 p1-Halo-pET29b。
所有克隆使用诺唯赞 ClonExpress II 一步克隆法完成。引物序列见附表 3。
体外蛋白表达与纯化
所有蛋白在大肠杆菌 Rosetta 菌株中表达,Ni-NTA 亲和纯化。0.4 mM IPTG 15℃ 诱导过夜。菌体超声破碎,离心取上清,Ni-NTA 结合,洗涤,咪唑洗脱。凝胶过滤色谱(Superdex-200)进一步纯化,保存于 4℃。
体积排阻色谱 —多角度激光联用(SEC-MALS)
使用 Superdex-200 柱,Wyatt DAWN HELEOS 系统检测,ASTRA 软件分析。
动态光散射( DLS)
使用 Wyatt DynaPro NanoStar,蛋白浓度 2 mg/mL,数据用 DYNAMICS 软件累积量拟合分析。
酵母转化与异源表达
质粒 NotI 线性化,转化裂殖酵母 LD328。42℃ 热激 40 分钟,EMM 培养基筛选,单菌落用于成像。
细胞与组织荧光成像
5 天龄拟南芥或烟草叶片用甘露醇、PEG、EG、重水等处理。根尖或叶盘制片,Zeiss LSM880 或 Nikon AXR 共聚焦显微镜成像。种子发育与萌发过程取样,剥去种皮后成像。酵母细胞用山梨醇或 NaCl 处理后成像。温度控制实验使用温控培养装置。
体外相分离实验
TEV 酶切去除 MBP 标签,蛋白稀释至所需浓度,加入 PEG、葡聚糖、聚蔗糖等,384 孔低吸附板成像,ImageJ 定量液滴大小、数量、荧光强度。
电场检测
使用 Di-4-ANEPPS 探针,共聚焦采集 535–545 nm 与 610–640 nm 荧光,计算比值反映局部电场强度。
微极性检测
蛋白与 Halo-SBD 按摩尔比 1:1 标记,脱盐,TEV 切标签,加 6% PEG 相分离。FLIM 荧光寿命成像,寿命越短极性越高。
荧光寿命成像 —荧光共振能量转移(FLIM-FRET)
Leica TCS SP8 显微镜,TCSPC 采集光子事件,双指数拟合计算平均寿命,分析分子间相互作用。
SAM8 进化分析
在 OrthoDB 数据库获取同源基因,共 143 个同源蛋白,选取 29 个代表性物种构建邻接树。无序区使用 IUPred2A 预测。
光漂白后荧光恢复( FRAP)
Nikon AXR 显微镜,488 nm 激光漂白,每秒采集一次,共 68 秒,ImageJ 分析荧光强度。
浊度检测
蛋白样品 96 孔板,VARIOSKAN FLASH 检测 600 nm 吸光度。
实时荧光定量 PCR(qRT-PCR)
TRIzol 提取 RNA,DNase I 消化,M-MLV 反转录,qPCR 检测,Actin 或 UBC 为内参。
渗透势测定
蒸汽压渗透压计 Vapro 5600 测定,水势 = 渗透压 × R × T。
免疫荧光染色
根尖 4% 多聚甲醛固定,纤维素酶消化,anti-Flag 一抗,Alexa Fluor 488 二抗,DAPI 染色,共聚焦成像。
RNA 荧光原位杂交(FISH)
5′-Cy5-dT35 探针检测 polyA mRNA,脱水、通透、杂交、洗涤、DAPI 染色,共聚焦成像。
质谱与互作分析
蛋白 SDS-PAGE 分离,胶内酶解,LTQ Orbitrap Elite 质谱检测,MASCOT 搜库,无标定量,筛选显著富集蛋白。
免疫共沉淀( Co-IP)
烟草叶片共表达 SAM8-GFP 与 Flag 标签蛋白,GFP 纳米磁珠富集,Western blot 检测。
双分子荧光互补( BiFC)
农杆菌注射烟草, 2 天后共聚焦显微镜检测荧光。
翻译组测序( Ribo-seq)与转录组测序(RNA-seq)
10 天龄幼苗 ±甘露醇处理 1 小时,分别建库测序。RNA-seq 纯化 polyA mRNA,Ribo-seq 富集核糖体保护片段(RPFs),去除 rRNA,建库测序。序列比对至 TAIR10,计算翻译效率 TE = RPKM(Ribo-seq)/ RPKM(RNA-seq)。差异基因与 GO 富集分析使用 clusterProfiler。
多聚核糖体图谱分析
5 天龄幼苗 ±甘露醇处理 1 小时,提取多聚核糖体,10–50% 蔗糖密度梯度离心,收集多糖组分,提取 RNA,qPCR 计算翻译效率。
转载本文请联系原作者获取授权,同时请注明本文来自孙学军科学网博客。 链接地址: https://blog.sciencenet.cn/blog-41174-1537074.html
上一篇: 热量限制降低补体C3和抗衰老有关 下一篇: 史上最长寿老人的秘密 欢迎参加科学网十佳博文评选活动! 主办单位: 支持单位: